Estudio de la variación secular en la talla de poblaciones humanas del centro de Argentina durante el Holoceno tardío [1]
Study of secular variation in the height of human populations from the center of Argentina during the late Holocene
Estudio de la variación secular en la talla de poblaciones humanas del centro de Argentina durante el Holoceno tardío [1]
Revista Jangwa Pana, vol. 18, núm. 3, pp. 396-419, 2019
Universidad del Magdalena

Esta obra está bajo una Licencia Creative Commons Atribución-NoComercial-CompartirIgual 4.0 Internacional.
Recepción: 12 Febrero 2019
Aprobación: 17 Junio 2019
Resumen: El objetivo de este trabajo es estudiar la variación secular en la talla de las poblaciones humanas que habitaron el centro de Argentina a lo largo del Holoceno tardío. Se espera encontrar una variación en la estatura y una disminución en el dimorfismo sexual en talla, asociado a los cambios en los modos de vida y las estrategias de subsistencia. Esto último relacionado con la transición de la caza y la recolección a una economía de tipo mixta, que complementa a la primera con la producción de alimentos. La muestra está compuesta por 42 individuos (18 femeninos, 24 masculinos), todos con información radiocarbónica asociada mediante AMS, que los sitúa entre 4058-387 14C años AP. Se registraron estaturas promedio altas para individuos femeninos (163,4 cm para fémur) y medianas para masculinos (168,3 cm para fémur). Los resultados, contrarios a las expectativas iniciales, sugieren un aumento del dimorfismo sexual a lo largo del tiempo, producto de una tendencia secular negativa de la talla en individuos femeninos y una leve tendencia al incremento de la estatura en masculinos. Las tendencias observadas en distintas regiones y por sexo no pueden ser explicadas atendiendo a un único factor explicativo. Más bien, la conjunción de causas ambientales y genéticas pueden estar interviniendo en los procesos que modelan la talla de los individuos a lo largo del tiempo.
Palabras clave: tendencia secular, dimorfismo sexual, clima, modos de vida, acervo genético.
Abstract: The main purpose of this work is to study the secular variation in stature of Late Holocene human populations that inhabited the centre of Argentina. It is expected to find a variation in height and a decrease in sexual dimorphism throughout the Holocene, associated with changes in lifestyle and subsistence practices, related to the transition from hunting and gathering lifestyle to a mixed economy, that complement hunt and gatherer with de food production. The sample is composed of 42 individuals (18 females, 24 males), all with associated radiocarbon information through AMS that places them between 4058-387 14C years BP. The results show high average statures for females (163.4 cm for the femur) and medium for men (168.3 cm for the femur). Contrary to the initial expectations, an increase in sexual dimorphism over time is suggested, as a result of a negative secular tendency in women and with a slight increase tendency in height among men. The observed trends, in different regions and by sex, can not be explained according to a single explanatory factor. Rather, the conjunction of environmental and genetic causes may intervene in the processes that model the size of individuals, over time.
Keywords: Secular trend, sexual dimorphism, climate, lifestyle, genetic pool.
Introducción
La estatura es una característica biológica fundamental de los individuos y de las poblaciones, valiosa para abordar diferentes problemáticas, desde aquellas vinculadas con el estudio de la salud y la nutrición, hasta procesos evolutivos a largo plazo (Pomeroy y Stock, 2012). El proceso que resulta en un cambio en las medidas corporales de una población en el transcurso del tiempo es conocido como tendencia secular en la talla (Millán et al., 2013, p. 422). Diversos estudios han demostrado que las variaciones de dichas medidas corporales representan un indicador válido del estado nutricional de un individuo o una población, y reflejan las condiciones socioeconómicas, políticas y ambientales en las que se desenvuelven las poblaciones (Komlos, 1998). Así, el mejoramiento en las condiciones contextuales puede traducirse en un incremento secular de la estatura promedio de individuos (Neves y Costa, 1998; Shin et al., 2012).
Durante las dos primeras décadas de vida, las actividades dominantes del organismo son el crecimiento y el desarrollo, fenómenos biológicos diferenciados y paralelos. Las respuestas adaptativas que comprometen o modifican el crecimiento y el desarrollo dependen de la etapa de la ontogenia en que se ven afectados y de la duración del episodio estresante, donde varios factores son capaces de generar modificaciones en la talla, como, por ejemplo, genéticos, endocrinos y ambientales. Pese a verse afectada por diversidad de factores, la estatura es una de las características de mayor heredabilidad en humanos: entre un 70 % y 90 % (Kemkes- Grottenhaler et al., 2005; Gustafsson et al., 2007). Sobre ella existe un control poligénico y varía considerablemente de una población a otra.
La estimación de la estatura a nivel individual es considerada un indicador de historia de vida, pues refleja el crecimiento a largo plazo. A nivel poblacional, la estatura promedio se transforma en un indicador sensible de la calidad de vida, un barómetro de la salud y la nutrición del grupo, que refleja las condiciones materiales de la sociedad en la que se desarrollan (Bogin, 1999). El estudio de la variación secular de la talla de los diversos individuos que han habitado la región de Sierras Centrales durante el Holoceno tardío, se constituye como una línea metodológica útil para discutir la importancia de aspectos relacionados con el impacto que produjeron determinadas estrategias de subsistencia adoptadas; así como el que produjo en la salud de las poblaciones la incorporación de nuevos recursos alimenticios a la dieta, a mediados del primer milenio de la era cristiana; y las migraciones humanas, en su escala temporal y espacial, entre otras cuestiones.
En la región estudiada se plantea que los procesos de intensificación y diversificación, manifestados en elementos materiales, de organización social y económica a partir de la transición hacia un modo de vida productor de alimentos alrededor de 1500 años AP, provocaron modificaciones en la dieta y en la actividad física de las poblaciones que habitaron las sierras y las llanuras del actual territorio de la provincia de Córdoba (Fabra et al., 2012, 2014, 2015). El aumento demográfico y la presión ambiental estimados para momentos previos a la conquista española habrían provocado episodios de estrés nutricional, mayor tensión social a nivel intra e interpoblacional, y emergencia de enfermedades (Laguens y Bonnin, 2009; Fabra et al., 2012, 2015). Estos procesos habrían impactado en el estado nutricional de los individuos y por ende en la talla de las poblaciones. De acuerdo con esto, se espera encontrar una variación en la estatura y una disminución en el dimorfismo sexual en talla a lo largo del Holoceno, asociadas a estos procesos socioeconómicos.
Las investigaciones bioarqueológicas de la antropología dental han propuesto que la moderada frecuencia de patologías de origen infeccioso y metabólico observada en la región durante el Holoceno tardío, concuerda con los valores esperados para poblaciones con economía cazadora-recolectora y de carácter mixto (Gonzalez, 2016; Fabra y Gonzalez, 2015). Los estudios de isotopos estables del carbono 13 sobre colágeno (13Ccol) sugieren que en el periodo posterior a 1200 años AP se estarían consumiendo alimentos producidos (Laguens et al., 2009; González, 2016). Esto, junto a una serie de cambios, como los fueron del tipo ambiental, habrían afectado las actividades cotidianas realizadas por las personas y, por ende, habrían generado modificaciones en el modo de vida de los individuos, observándose un leve deterioro en la salud para épocas tardías (Laguens y Bonnin, 2009; González, 2016). La relación entre un desmejoramiento de la salud y la transición hacia una economía de carácter agrícola fue abordada por Cohen y Armelagos (1984) y Roosevelt (1984), quienes asociaron la reducción de la edad promedio de muerte y la alta frecuencia de lesiones esqueletales al surgimiento de las prácticas agrícolas junto a la sedentarización.
Los estudios sobre cambios degenerativos y entesiales en poblaciones de la misma región sugieren que para momentos del Holoceno tardío final disminuyen las frecuencias de modificaciones óseas en miembros inferiores y columna, a la vez que la prevalencia de cambios entesiales es mayor en miembros superiores, lo que sugiere una disminución en la movilidad residencial (Salega, 2016; Salega y Fabra, 2013).
Las poblaciones que habitaron la región serrana y las llanuras orientales del centro de la Argentina, en la actual provincia de Córdoba, han experimentado diversos cambios y transformaciones en sus modos de vida a lo largo del Holoceno tardío. Las investigaciones arqueológicas (Laguens, 1999; Bonifliogio, 2009;Laguens et al., 2009; Laguens y Bonnin, 2009; Medina et al., 2016), bioarqueológicas (Fabra et al., 2012, 2014; Fabra y González, 2015; Salega y Fabra, 2013) y genéticas (Nores y Demarchi, 2011; Nores et al., 2011) sugieren que existen diferencias entre ambas regiones en cuanto a la cultura material y las estrategias de subsistencia desarrolladas en cada una a lo largo del tiempo, el tratamiento del cuerpo en contextos mortuorios (Fabra et al., 2009), la variabilidad poblacional a partir de la variación morfológica craneofacial (Fabra, 2014) y el ADN antiguo (Nores y Demarchi, 2011; Nores et al., 2011, 2017).
En los últimos años se han realizado investigaciones que abordan el estudio de la estructura y la historia biológica de las poblaciones que habitaron (Fabra, 2009, 2014; Fabra y Demarchi, 2009, 2012, 2013; Nores y Demarchi, 2011; Nores et al., 2011, 2017) y habitan (García y Demarchi, 2006; Pauro et al., 2010) el actual territorio de la provincia de Córdoba. El análisis de la variabilidad de rasgos epigenéticos y morfogeométricos craneofaciales muestra tendencias hacia un agrupamiento geográfico entre las poblaciones asentadas en los diferentes valles serranos, por un lado, y las que ocuparon los ambientes de llanuras (Fabra et al., 2005; Fabra y Demarchi, 2011), por el otro.
Investigaciones sobre ADN mitocondrial realizados a partir de un conjunto de muestras arqueológicas de distintas regiones de Córdoba sugieren que hace 1200 años AP habría ocurrido una diferenciación genética entre las poblaciones que habitaron distintos espacios geográficos de la provincia de Córdoba (Nores y Demarchi, 2011; Nores et al., 2011; Fabra et al., 2014). Sin embargo, estas variaciones observadas en los haplogrupos no implicarían necesariamente la configuración de dos poblaciones distintas en estos espacios geográficos, ya que existe continuidad genética a lo largo del tiempo. García y colaboradores (2012) plantean la existencia de una antigua metapoblación en las Sierras Pampeanas, de la cual Córdoba formaría parte, que se habría caracterizado por su alta frecuencia y la diversidad interna del haplogrupo D1j, cuyo origen habría estado en esta región. Se debe destacar que, a lo largo del tiempo, las relaciones biológicas con poblaciones vecinas debieron producir modificaciones en el acervo genético y, seguramente, en las prácticas culturales y en las formas de conocer e interpretar el mundo.
Como hipótesis general se plantea que el proceso de intensificación y diversificación de la organización social y económica a partir de la transición hacia un modo de vida productor de alimentos entre 2000-1500 años AP (Laguens y Bonnin, 2009; Fabra et al., 2012, 2015), habría provocado modificaciones en la dieta y en la actividad física de las poblaciones que habitaron las sierras y las llanuras del actual territorio de la provincia de Córdoba, lo que resultó en un leve deterioro del estado general de la salud, en comparación con las condiciones de vida basadas en la caza y la recolección. Además, el aumento demográfico y la presión ambiental estimados para momentos previos a la conquista española habrían provocado episodios de estrés nutricional, mayor tensión social a nivel intra e interpoblacional, y emergencia de enfermedades. Estos procesos tendrían impacto en el estado nutricional de los individuos como en sus actividades cotidianas y, por ende, en la expresión de la estatura de las poblaciones. Entonces, en esta investigación se espera encontrar decrecimiento en la talla y una disminución en el dimorfismo sexual en talla a lo largo del Holoceno y entre regiones, asociado a estos procesos socioeconómicos.
Materiales y métodos
Muestra
La muestra está compuesta por un total de 42 individuos (18 femeninos y 24 masculinos), que proceden de diversos sitios arqueológicos ubicados tanto en la zona de las sierras ―vertientes oriental y occidental― como en las llanuras extraserrana oriental. Fueron seleccionados por su buen estado de conservación, con presencia de miembros inferiores, y por poseer información contextual y cronológica absoluta. Esto permitió situar los resultados en un marco temporal y espacial concreto, entre los 4058± 89 y los 387± 41 14C años AP (Tabla 1, Figura 1).
| Nombre - Sitio | Edad 14C AP (sin calibrar) | Sexo | Edad | Región | Tipo de dieta | Long. máx fémur (cm) | Long. máx. tibia (cm) | Estatura (cm) | Referencias |
| 2012 Ea La Elisa I2 | 4058± 89 | M | AMD | Llanuras | Mixta | 43,6 cm | 43,7 cm | 166,5 cm | Gonzalez y Fabra (2018) |
| 123 ED010 Cuad R | 2562± 47 | F | AJ | Llanuras | Mixta | 42 cm | 36,1 cm | 162,8 cm | Fabra et al. (2014) |
| 122 ED010 Cuad O | 2438± 47 | F | AMD | Llanuras | C-R | 43,6 cm 43,6 cm | - | 166,3 cm | Fabra et al. (2014) |
| 121 ED010 Cuad Q | 2400± 47 | F | AM | Llanuras | C-R | 41,5 cm | 35,7 cm | 161,7 cm | Fabra et al. (2014) |
| 126 ED010 Cuad S | 2331± 46 | M | AMD | Llanuras | Mixta | 46 cm | - | 171,4 cm | Fabra et al. (2014) |
| Emist06I | 2282± 47 | F | AMD | Llanuras | Mixta | 43,5 cm | 36,8 cm | 166 cm | Gonzalez y Fabra (2018) |
| El Vado (Dto. Punilla, Cba) | 2156± 86 | F | AMD | Sierras | C-R | 41,7 cm | 35 cm | 162,2 cm | Fabra et al (2012) |
| El Diquecito 08, Cuad, G, Ind.1 | 1911 ± 59 | M | AMD | Llanuras | Mixta | 42,5 cm | 36,3 cm | 163,9 cm | Fabra et al (2012) |
| 2009 Ea La Elisa I1 | 1890± 49 | M | AMD | Llanuras | Mixta | 44 cm | 37,3 cm | 167,1 cm | Gonzalez y Fabra (2018) |
| 2009 ECO 09 I1 –Sitio Ecoterra | 1881± 39 | M | AJ | Sierras | Mixta | 43 cm | 36,1 cm | 165,1 cm | Gonzalez y Fabra (2018) |
| 94A - Colonia Muller site | 1585± 15 | F | AJ | Llanuras | Mixta | - | 37,1 cm | 170,2 cm | Fabra y Gonzalez (2008), Fabra et al. (2009) |
| 67, La Granja Site, ("I1) | 1280± 20 | F | AMD | Sierras | Mixta | 44,9 cm | 38,4 cm | 169,2 cm | Laguens et al. (2007); Fabra et al.(2009) |
| LagPlata01 | 1241± 57 | M | AMD | Llanuras | Mixta | 43,8 cm | 37,8 cm | 166,8 cm | Gonzalez y Fabra (2018) |
| El Diquecito 08, Cuad. E, Ind.1 | 1192 ± 40 | M | AMD | Llanuras | Mixta | 42,3 cm | 36,4 cm | 163,5 cm | Fabra et al. (2012) |
| 2011 Potrerillo de Larreta | 1067± 77 | M | AJ | Sierras | Mixta | 42,6cm | - | 164,1 cm | Gonzalez y Fabra (2018) |
| 87A - Orihuela site (1) | 1045± 15 | F | AJ | Llanuras | Mixta | 40,4 cm | 34,2 cm | 159,5 cm | Fabra et al. (2009) |
| 95A - San Esteban site | 965± 15 | F | AJ | Sierras | Mixta | 42 cm | - | 162,9 cm | Fabra y Gonzalez (2008), Fabra et al. (2009) |
| Loma Bola- CB (Dto. San Javier) | 954± 85 | F | AMD | Sierras | Mixta | - | 34,1 cm | 162,9 cm | Fabra et al. (2012) |
| Loma Bola- CA2 (Dto. San Javier) | 954± 85 | F | AJ | Sierras | Mixta | 40,7 cm | - | 160 cm | Fabra et al. (2012) |
| Loma Bola- CA (Dto. San Javier) | 954± 85 | M | AMD | Sierras | Mixta | 42,4 cm | 34,3 cm | 163,8 cm | Fabra et al. (2012) |
| El Diquecito 08, Cuad. L, Ind.1 | 937± 150 | F | AJ | Llanuras | Mixta | 41 cm | 35 cm | 160,7 cm | Fabra et al. (2012) |
| Guasmara Site, ("I1") | 920± 20 | M | AJ | Sierras | Mixta | 45,2 cm | 38,5 cm | 169,8 cm | Laguens et al. (2007), Fabra y Gonzalez (2008), Fabra (2008), Fabra et al (2009) |
| 66B, Guasmara Site, ("I2) | 920± 20 | M | AJ | Sierras | Mixta | 46,7 cm | 39,1 cm | 173 cm | Laguens et al. (2007), Fabra y Gonzalez (2008) |
| E6/56 | 881± 150 | M | AM | Sierras | Mixta | 45,5 cm | 38,9 cm | 170,3 cm | Fabra et al. (2012) |
| 127 Isla Orihuela I2 | 761± 41 | M | AMD | Llanuras | Mixta | 44 cm | 35,8 cm | 167,2 cm | Fabra et al. (2012) |
| El Diquecito 08, Cuad. B, Ind 1 | 750± 85 | M | AJ | Llanuras | Mixta | 44,6 cm | 36,4 cm | 168,5 cm | Fabra et al. (2012) |
| Rosca Yaco, I1 | 705± 131 | M | AMD | Sierras | Mixta | 43,6 cm | - | 166,3 cm | Fabra et al. (2012) |
| 124 ED08 Cuad A | 698± 42 | F | AMD | Llanuras | Mixta | 41,2 cm | 33,9 cm | 161,2 cm | Fabra et al.(2012) |
| 70A - Banda Meridional del Lago site | 695± 20 | F | AMD | Sierras | Mixta | - | 34,1 cm | 163 cm | Fabra y Gonzalez (2008), Fabra et al. (2009) |
| Copina I2 | 680± 40 | M | AMD | Sierras | Mixta | 42,3 cm | 33,6 cm | 163,6 cm | Fabra et al. (2012) |
| La Orihuela (Orih08) | 664± 33 | M | AJ | Llanuras | Mixta | 44,6 cm | - | 168,5 cm | Este trabajo |
| CO 09 I1 -Constantinopla | 619± 43 | M | AJ | Sierras | Mixta | 42,7 cm | - | 164,3 cm | Gonzalez y Fabra (2018) |
| 17B, Ayampitin Site, (I1) | 600± 20 | M | AJ | Sierras | Mixta | 47 cm | - | 173,7 cm | Laguens et al. (2007), Fabra (2008) |
| 125 ED010 Cuad P | 597± 41 | F | AMD | Llanuras | Mixta | - | 36,2 cm | 168 cm | Fabra et al. (2014) |
| El Diquecito 08, Cuad. M, Ind. 1 | 537± 57 | F | AMD | Llanuras | Mixta | 41 cm | 33,7 cm | 160,8 cm | Fabra et al.(2012) |
| 2011 Lote 5 Sta Rosa CI I2 | 533± 42 | F | AM | Llanuras | Mixta | 42,5 cm | 35,8 cm | 163,9 cm | Gonzalez y Fabra (2018) |
| 2011 Lote 5 Sta Rosa CI I1 | 533± 42 | F | AMD | Llanuras | Mixta | 39,6 cm | 33,7 cm | 157,8 cm | Gonzalez y Fabra (2018) |
| Rincon Site (RI-S2) | 520± 15 | M | AMD | Sierras | Mixta | 46,2 cm | 38 cm | 171,9 cm | Laguens et al. (2007) |
| 2011 Isla Orihuela. PS, I1 | 487± 45 | M | AJ | Llanuras | Mixta | 43 cm | 35,7 cm | 165 cm | Gonzalez y Fabra (2018) |
| I2 Cañada Larga –Mina Clavero | 481± 57 | M | AMD | Sierras | Mixta | 46,6 cm | 38,1 cm | 172,7 cm | Este trabajo |
| Ischilin I1 | 459± 40 | M | AJ | Sierras | Mixta | - | 39,1 cm | 175 cm | Fabra et al. (2012) |
| Nunsacat (Copacabana, Cba). Individuo 2 | 387± 41 | M | AMD | Sierras | Mixta | 46,5 cm | 39,2 cm | 172,5 cm | Fabra et al. (2012) |
Referencia: F (Femenino), M (Masculino), C-R (Cazador Recolector), A (Adulto, 20+ años), AJ (Adulto Joven, 20-34 años), AMD (Adulto Medio, 35-49 años), AM (Adulto Mayor, 50+ años).
Fuente: elaboración propia.
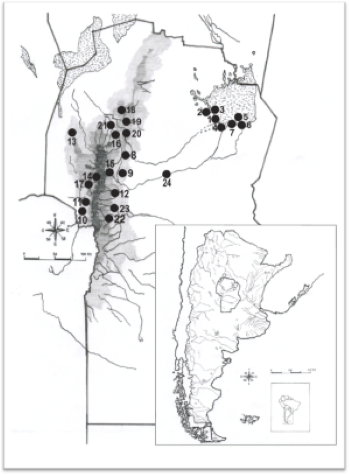
Figura 1.
Procedencia de las muestras relevadas en el presente estudio
1. Sitio El Diquecito (11 individuos) 2. Sitio Estancia La Elisa (2 individuos) 3. Sitio El Mistolar 4. Sitio Laguna del Plata, 5. Sitio Isla Orihuela (2 individuos), 6. Sitio La Orihuela (2 individuos), 7. Sitio Colonia Muller, 8. Sitio Ecoterra, 9. Sitio Potrerillo de Larreta, 10. Sitio Loma Bola (3 individuos), 11. Sitio Guasmara (2 individuos), 12. Sitio Los Molinos, 13. Sitio Rosca Yasco, 14. Sitio Copina, 15. Sitio Constantinopla, 16. Sitio Ayampitin, 17. Sitio Cañada Larga, 18. Sitio Ischilin y Nunsacat, 19. Sitio El Vado, 20. Sitio La Granja, 21. Sitio San Esteban, 22. Sitio Banda Meridional del Lago, 23. Sitio Lote 5 Santa Rosa (2 individuos), 24. Sitio Rincón.
Fuente: elaboración propia.Metodología
Se tuvieron en cuenta los procedimientos establecidos por Buikstra y Ubelaker (1994) para la determinación del sexo y la estimación de edad aproximada al momento de muerte en cada uno de los individuos analizados. Para la determinación del sexo se consideraron las características morfológicas de la pelvis (región subpúbica, escotadura ciática y surco preauricular) y del cráneo (desarrollo cresta nucal, tamaño del proceso mastoideo, ángulo del margen supraorbital, prominencia de la glabela y proyección de la eminencia mental). En caso de ausencia o mala preservación del cráneo o pelvis, el sexo se estimó a partir de funciones discriminantes para el fémur, desarrolladas para poblaciones del sur de Sudamérica (Beguelin y González, 2008). Para la estimación de edad, se siguió la metodología compilada por Buikstra y Ubelaker (1994), que considera rasgos cualitativos del cráneo (desarrollo dental como cierre de las suturas craneales), pelvis (sínfisis púbica y superficie auricular del ilion) y costillas (metamorfosis de los fines esternales de costillas). En los casos en que no se pudo estimar la edad aproximada del individuo al momento de muerte, se procedió a analizar aquellos fémures y tibias que se encontraban totalmente fusionados en todos sus elementos anatómicos (Scheuer y Black, 2000).
Estimación de la talla en poblaciones arqueológicas
Fémur y tibia son los elementos óseos que aportan la mejor estimación de la estatura final en vida (Ruff et al., 1991; Beguelin, 2009, 2010). El criterio de selección de los miembros inferiores para la estimación de talla se fundamenta en que se ha observado que las poblaciones sometidas a situaciones de estrés crónico (al menos en las primeras etapas de vida) presentan una reducción de la estatura total, a expensas del acortamiento de los miembros inferiores. Por ello, en casos donde se dan disrupciones de la homeostasis, procesos de desnutrición crónica o condiciones ambientales desfavorables, el crecimiento del tronco ―y, en mayor magnitud, la longitud de las piernas― se ve afectado por este tipo de procesos (Tanner, 1962; Van Wieringen, 1986).
Para llevar adelante las mediciones de los miembros inferiores se utilizaron los criterios y las recomendaciones establecidas por Buikstra y Ubelaker (1994): se registraron las longitudes máximas para fémur y tibia para la estimación de la talla. Para el registro y la toma de medidas, se utilizó el calibre digital Mitutoyo (0.001 mm de precisión) y la tabla osteométrica (0.01 cm de precisión). Para el caso de restos óseos fragmentados, se utilizó el método y los criterios establecidos por Steele y Mckern (1969), los cuales permitieron realizar el cálculo aproximado de la longitud máxima de fémur y tibia.
Una vez tomadas las mediciones de longitud máxima para fémur y tibia, se procedió a estimar la estatura, considerando las fórmulas para grupos Mongoloides de Trotter y Gleser (1958):
Estimación por fémur: 2,15 x Long. Máx. Fémur (cm) + 72,57.
Estimación por tibia: 2,39 x Long. Máx. Tibia (cm) + 81,45.
Para las estimaciones realizadas, siempre se priorizo el fémur sobre la tibia y el lado derecho sobre el izquierdo, ya que proporcionan una mejor estimación de la talla (Formicola y Giannecchini, 1999). En el caso de la ausencia de alguno de los elementos anatómicos mencionados, se procedió a reemplazarlo por el que se encontrara presente.
En la toma de medidas y en el análisis de los datos puede haber inconsistencias o errores en la medición, resultado de una incorrecta definición o medición de los puntos (Krenzer, 2006). Para reducir el rango de error intraobservador, se tomaron medidas sobre un total de 42 individuos, repitiendo la medición a los 15 días. No se observaron diferencias estadísticamente significativas entre las mediciones.
A lo largo del estudio se realizaron diversos agrupamientos: para evaluar el dimorfismo sexual se tuvo en cuenta el sexo de los individuos (femenino o masculino); para evaluar la tendencia secular de la talla se tuvo en cuenta la cronología de las muestras (periodo previo y posterior a 1200 años AP), los modos de vida con base en los isótopos estables o la cronología, en especial los de 13Ccol (Coltrain y Leavitt, 2002) (caza y recolección, o dietas mixtas que integran caza y recolección con producción de alimentos) y las regiones geográfico-culturales de procedencia (sierras y llanuras). Vale aclarar que la cronología propuesta de 1200 años AP como corte para las comparaciones se funda en investigaciones genéticas de la región, que dan cuenta de un cambio en el acervo genético de las poblaciones, que coincidiría, en cierta forma, con la incorporación de prácticas hortícolas (Nores et al., 2011).
Análisis estadísticos
En función de los objetivos propuestos en el presente trabajo, se llevaron adelante los siguientes análisis estadísticos, siguiendo el diseño de investigación propuesto por Millán y colaboradores (2013):
- 1. Para evaluar la presencia de asimetría bilateral, es decir, si había diferencias estadísticamente significativas entre las medidas de cada individuo para la lateralidad derecha e izquierda, se compararon las estaturas calculadas a partir de fémur y tibia, considerando la lateralidad y el sexo. Esto utilizando la prueba t de Student (muestras apareadas) con un valor de p de 0,05.
- 2. Luego, se compararon las estaturas estimadas considerando las regiones (sierras, llanuras), los modos de vida (cazador-recolector, economía mixta) y periodos (pre 1200 AP, post 1200 AP). Se utilizó en este caso la prueba U de Mann-Whitney (muestras independientes) con un valor de p de 0,05.
- 3. Finalmente, se estimó el dimorfismo sexual con la prueba de Mann-Whitney. Esta prueba es el método más adecuado para comparar poblaciones cuyas distribuciones no son normales.
Resultados
Considerando el total de la muestra (Tabla 1), se clasifican los 18 individuos femeninos relevados entre los “Hipsisomos Altos” (entre 159,3- 167,9 cm) a partir de la media de 164 cm, con alturas máximas de 170,2 cm y mínimas de 157,8, siguiendo las clases estaturales propuestas por Martin y Saller (1957). Para el caso de los 24 individuos masculinos analizados, se ubican dentro de los “Mesosomos Supermedianos” (entre 167- 169,9 cm) a partir de la media de 169,2 cm, con tallas máximas de 175 cm y mínimas de 163,5 cm. Se observó que, en promedio, en las tallas estimadas según la media, los individuos masculinos son 5,2 cm más altos que los femeninos. Para el caso de la estatura calculada a partir de fémur en los individuos femeninos, se observó una talla mínima de 157,6 cm y una altura máxima de 169,2 cm, con una media de 163,4 cm. Para el caso de las tibias, la talla mínima observada fue de 161,6 cm y una altura máxima de 173,2 cm, con una media de 167,4 cm. Vemos que las estaturas calculadas sobre ambos elementos se ubican en la clasificación “Hipsisomos Altos” (entre 159,3- 167,9 cm) propuesta por Martin y Saller (1957). En las estimaciones de estatura para individuos masculinos, la talla mínima calculada a partir de fémur fue de 162,9 cm y una altura máxima de 173,7 cm, con una media de 168,3 cm. En el caso de las tibias, la mínima fue de 163 cm y la máxima de 175 cm, con una media de 169 cm. Retomando la clasificación de Martin y Saller, las estaturas, si se consideran ambos elementos, se mantienen dentro de los “Mesosomos Supermedianos” (entre 167- 169,9 cm).
Lateralidad
En la Tabla 2 se presentan los resultados de la comparación de las estaturas obtenidas con los elementos anatómicos de distinta lateralidad, por sexo. De las cuatro comparaciones realizadas, los resultados fueron significativos en el caso de las estaturas estimadas con fémur en individuos femeninos, por lo que solo se tuvo en cuenta el fémur derecho para las estimaciones de la talla. Cuando se encontró ausente el fémur derecho, se asignó la estimación realizada con base en la tibia.
| Sexo | Variables | n | Media | Desviación tip. | Error tip. de la media | t | gl | Sig. (Bilateral) |
| Masculino | Talla fémur izq./ talla fémur der. | 13 | 0,315 | 0,722 | 0,200 | 1,575 | 12 | 0,141 |
| Masculino | Talla tibia izq./ talla tibia der. | 11 | 0,700 | 2,248 | 0,677 | 1,033 | 10 | 0,326 |
| Femenino | Talla fémur izq./ talla fémur der. | 7 | 0,471 | 0,509 | 0,192 | 2,451 | 6 | 0,050 |
| Femenino | Talla tibia izq./ talla tibia der. | 6 | -0,433 | 0,564 | 0,230 | -1,880 | 5 | 0,119 |
Referencias: n (número de individuos), desviación tip. (desviación típica), error tip. de la media (error típico de la media), t (valor de la prueba t), gl (grados de libertad), sig (bilateral) (significación bilateral).
Fuente: elaboración propiaDimorfismo sexual
Para determinar si existió dimorfismo sexual en la muestra analizada, se aplicó la prueba de Mann- Whitney para las distintas comparaciones ya nombradas. Se encontraron diferencias significativas en y entre periodos cronológicos. En la Tabla 3 se presentan los resultados de las comparaciones realizadas con base en el sexo entre los periodos cronológicos propuestos. En el caso de los individuos masculinos previos al 1200 AP no se encontraron diferencias significativas para fémur al comparar con femeninos de ambos periodos, pero sí para la comparación con tibia. Tomando al grupo de individuos masculinos posteriores al 1200 AP se encontraron diferencias significativas para tibia en comparación con los femeninos de momentos iniciales al Holoceno tardío, pero no para fémur. En la comparación entre sexos del mismo periodo (post 1200 AP) sí se encontraron diferencias significativas tanto para fémur como para tibia.
Centrando la atención en los resultados obtenidos entre periodos, y en particular aquellos que resultaron significativos, se puede destacar:
Entre los individuos femeninos de ambos periodos existe una estatura de 163,76 cm en momentos previos a los 1200 años AP y, para posteriores, 160,55 cm (considerando fémur derecho). Esto da una diferencia de 3,21 cm entre las medias. En la comparación de tibias, para momentos iniciales del Holoceno tardío la estatura media es de 168,7 cm y para momentos finales es de 163,82 cm. Esto da una diferencia de 4,88 cm.
En el caso de los individuos masculinos y femeninos de momentos tardíos se observaron diferencias significativas en las estaturas estimadas con fémures y con tibias.
| Femeninos PRE 1200 | Femeninos POST 1200 | Masculinos PRE 1200 | Masculinos POST 1200 | |
| Femeninos PRE 1200 | - | Fémur/Tibia 0,05/0,08 | Fémur/Tibia NS/0,02 | Fémur/Tibia 0,001/0,001 |
| Femeninos POST 1200 | - | - | Fémur/Tibia NS/0,001 | Fémur/Tibia 0,001/0,000 |
| Masculinos PRE 1200 | - | - | - | NS |
Referencia: NS ( no significativo).
Fuente: elaboración propia.La media para masculinos es de 168,53 cm (fémur) y para femeninos de 160,55 cm (fémur), existiendo un dimorfismo sexual de 7,98 cm en este periodo. Para las estaturas estimadas considerando la tibia, en los masculinos la media es de 170,45 cm y en femeninos es de 163,82 cm. Se observa un margen de 6,63 cm entre ambas medias para el mismo periodo.
Variación regional en la estatura
En el caso de los individuos femeninos, la media calculada para fémures derechos fue de 161,8 cm en llanuras y de 160,9 cm en sierras, tendencia similar a la observada en las estaturas calculadas con base en la tibia, donde en la región llanuras se obtuvieron las tallas más elevadas. Las diferencias no fueron estadísticamente significativas (Tabla 4).
| Región | n | Media | DE | Min (cm) | Max (cm) | U | Sig. (2-tailed) | |
| Talla FD (long. máx) | Llanuras | 6 | 161,88 | 2,41 | 159,5 | 166,3 | ||
| Sierras | 4 | 160,97 | 2,65 | 157,8 | 163,9 | 10,00 | 0,76 | |
| Talla T (long. máx) | Llanuras | 9 | 165,86 | 3,26 | 161,6 | 170,2 | 28,50 | 0,75 |
| Sierras | 7 | 165,37 | 3,88 | 161,6 | 173,2 |
Referencias: FD (fémur derecho), T (tibia), DE (desvío estándar), U (valor p de la prueba U de Mann-Whitney)
Sig. (2-tailed) (significación)
Fuente: elaboración propiaPara los individuos masculinos, la media calculada en fémures fue de 167,5 cm en llanura, y de 168,2 cm para sierras. Tendencia similar se observó al estimar la estatura a partir de tibias, donde se obtuvieron tallas más elevadas para individuos procedentes de las sierras. Las diferencias no fueron estadísticamente significativas entre regiones (Tabla 5).
| Columna1 | Región | n | Media (cm) | DE | Min (cm) | Max (cm) | U | Sig. (2-tailed ) |
| Talla F (long máx) | Llanuras | 11 | 167,50 | 2,82 | 165 | 172,9 | 62,00 | 0,83 |
| Sierras | 12 | 168,25 | 4,10 | 163,8 | 173,7 | |||
| Talla T (long máx) | Llanuras | 9 | 169,74 | 2,51 | 166,9 | 173,7 | 28,00 | 0,29 |
| Sierras | 9 | 170,94 | 4,77 | 163 | 175,2 |
Referencias: FD (fémur derecho), T (tibia), DE (desvío estándar), U (valor p de la prueba U de Mann-Whitney)
Sig. (2-tailed) (significación).
Fuente: elaboración propia.Variación en la talla a lo largo del Holoceno
En la Figura 2 se presentan las estaturas estimadas por sexo, considerando la cronología de cada individuo. Podemos observar un aumento de la talla en los individuos masculinos en momentos tardíos. Para el caso de los femeninos, se observa un patrón opuesto, es decir, con disminución de la talla a lo largo del Holoceno
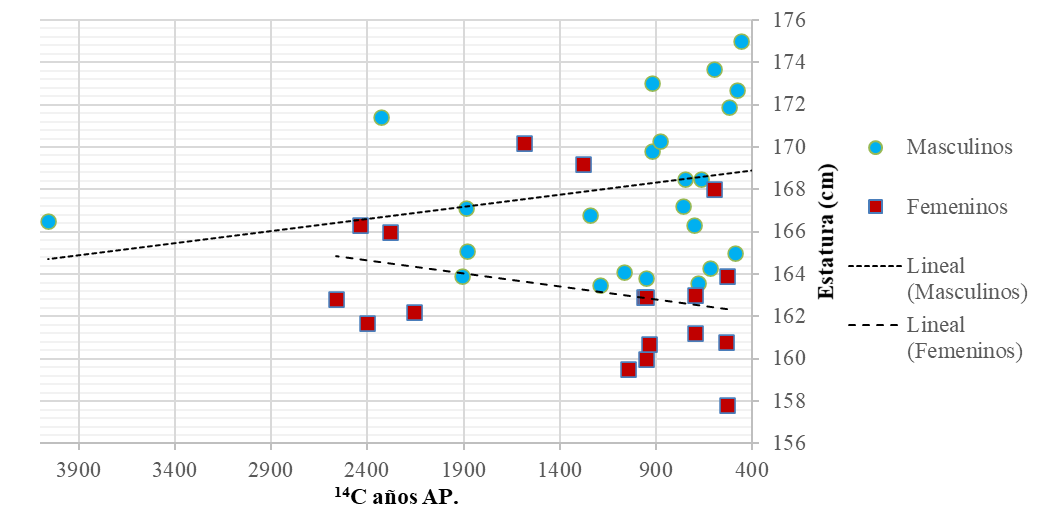
Figura 2.
Variación temporal de la estatura en la muestra analizada.
Referencias: Cuadrados para femeninos, círculos para masculinos, línea de tendencia con guiones para femeninos, línea de tendencia con puntos para masculinos.
Fuente: elaboración propiaVariación en la estatura considerando los modos de vida
Finalmente, el interés fue poner a prueba si los cambios en las distintas estrategias adaptativas que se desarrollaron en la región en los últimos 4000 años habían impactado en la talla de los individuos. Para ello, se agruparon los individuos en dos grandes categorías, caza-recolección y modo de vida mixto (caza, recolección y horticultura), teniendo en cuenta la información arqueológica e isotópica asociada a cada individuo y su cronología.
En el caso de los individuos femeninos, se observó la estatura estimada para fémur. En aquellos agrupados en un modo de vida basado en la caza y la recolección fue de 164,2 cm, mientras que la estatura promedio en individuos que habrían desarrollado estrategias mixtas seria de 160,8 cm. Un patrón similar se observó al estimar estaturas a partir de tibias, siendo más elevadas las estaturas para individuos con un modo de vida basado en la caza y la recolección. Pese a estas diferencias, las mismas no fueron estadísticamente significativas (Tabla 6). Finalmente, la estatura estimada para individuos masculinos bajo un modo de vida basado en la caza y la recolección es de 166,9 cm (fémur), mientras que para individuos que desarrollaron estrategias de subsistencia de tipo mixta es de 167,9 cm. Las estaturas estimadas a partir de la tibia fueron similares. No se encontraron diferencias significativas en el caso de las comparaciones realizadas tanto en fémur como en tibia (Tabla 7).
| Modo de vida | n | Media | DE | Min (cm) | Máx (cm) | U | Sig. (2-tailed) | |
| Talla FD (long. máx) | C-R | 2 | 164,25 | 2,89 | 162,2 | 166,3 | 2,00 | 0,17 |
| M | 8 | 160,83 | 1,89 | 157,8 | 163,9 | |||
| Talla T (long. máx) | C-R | 3 | 167 | 2,26 | 164,9 | 169,4 | 12,50 | 0,36 |
| M | 13 | 165,33 | 3,64 | 161,6 | 173,2 |
Referencias: FD (fémur derecho), T (tibia), C-R (caza y recolección), M (mixta), DE (desvío estándar), U (valor p de la prueba U de Mann-Whitney)
Sig. (2-tailed) (significación).
Fuente: elaboración propia.
| Modo de vida | n | Media | DE | Min (cm) | Máx (cm) | U | Sig. (2-tailed) | |
| Talla F (long. máx) | C-R | 5 | 166,90 | - | 164,1 | 173,1 | 36,00 | 0,51 |
| M | 15 | 167,94 | 3,56 | 163,9 | 173,7 | |||
| Talla T (long. máx) | C-R | 4 | 170,30 | - | 164 | 173,8 | 25,00 | 0,76 |
| M | 14 | 170,34 | 3,87 | 163 | 175,2 |
Referencias: FD (fémur derecho), T (tibia), C-R (caza y recolección), M (mixta), DE (desvío estándar), U (valor p de la prueba U de Mann-Whitney), Sig. (2-tailed) (significación).
Fuente: elaboración propia .Discusión y conclusiones
La presente investigación permitió confirmar que existen diferencias significativas en la talla entre períodos, que dan cuenta de dimorfismo sexual en la muestra analizada a lo largo del Holoceno tardío. Dado que la estatura de una población puede variar en relación con factores ambientales (i.e. clima, estatus socioeconómico, la alimentación) o biológicos (i.e. migraciones), interesa discutir estos factores a fin de interpretar cuáles pueden haber sido los causales que generaron estas variaciones a través del tiempo.
Dentro de los factores que pueden considerarse causales de cambios en la talla se encuentra el clima. Los cambios en las condiciones climáticas influyen directamente sobre la disponibilidad de los recursos y la calidad de la dieta, inciden en las condiciones ambientales donde los individuos crecen y se desarrollan, así como en las estrategias socioeconómicas adoptadas por las poblaciones. El estudio llevado adelante por Koepke (2016) reconoce que existe diversidad de factores que influyen en la estatura de las poblaciones, sugiriendo que los cambios climáticos afectan las estrategias de subsistencia y, colateralmente, la salud de estas, lo que afecta la calidad de vida y produce variación en la talla.
En Argentina, investigaciones realizadas sobre poblaciones que habitaron Patagonia los últimos 2000 años, han asociado la latitud y las condiciones climáticas a la talla y al tamaño corporal (Beguelin, 2010; Beguelin y Barrientos, 2005; Bernal et al., 2013). En esos trabajos se ha observado que los mayores tamaños están en los climas más fríos (Beguelin, 2011; Hernández et al., 1997). Este tipo de procesos puede ser explicado por la convergencia de diferentes factores y procesos, como la selección direccional de los caracteres, la adaptación climática local y los movimientos migratorios de pequeño y mediano rango (Beguelin y Barrientos, 2005). En lo que respecta al presente estudio, no se podrían realizar asociaciones entre clima y dispersión geográfica ya que se trabajó con muestras de dos regiones cercanas en términos geográficos, pero sí se pueden relacionar los cambios en la talla con la variación climática, considerando que hubo periodos que tuvieron efectos climáticos opuestos en la región, de alternancia de climas húmedos y templados con fríos y áridos.
En la región analizada en este trabajo, durante los últimos 5000 años se sucedieron una serie de cambios climáticos de magnitud: por un lado, la Anomalía Climática Medieval (ACM) que se extendió entre 1400-800 años AP (Piovano et al., 2009) y cuyos efectos en la región central de Argentina fueron las precipitaciones abundantes con aumento de las cuencas hídricas, así como temperaturas más benignas y humedad creciente, similares a las actuales. Esto favoreció el avance y la colonización del bosque Chaqueño ubicado en siglos anteriores varios cientos de kilómetros hacia el norte (Laguens y Bonnin, 2009). Por otro lado, la Pequeña Edad de Hielo (PEH) se extendió entre 600 y 200 años AP ―siglo XIV a mediados del XIX de nuestra era― (Piovano et al., 2009). Sus efectos en la región, opuestos a los de la ACM, implicaron la aridización y el descenso de la temperatura, promediando un clima frío donde, según las crónicas, eran frecuentes las nevadas, aunque sin nieves permanentes, junto a limitadas precipitaciones anuales con el retroceso de los cursos de agua (déficit hidrológico) y con un notable deterioro de los recursos potencialmente consumidos (Carignano, 1999).
Es interesante relacionar la variación en la talla con los procesos de cambio climático ocurridos en esta región, donde las tallas más elevadas se observan en momentos en que se desarrolló la ACM y sus condiciones climáticas más benignas, que habrían posibilitado una mayor disponibilidad de recursos por aproximadamente un lapso de 600 años (Carignano, 1999; Laguens y Bonnin, 2009). En momentos posteriores, la talla se mantiene con un leve incremento para los individuos masculinos, y un descenso significativo de la talla en el caso de las mujeres, en un período que coincide con el desmejoramiento climático ocurrido bajo la PEH.
Trabajos bioarqueológicos que han abordado cuestiones vinculadas con la salud oral y los marcadores de actividad física en esta región (Fabra et al., 2012, 2015; González y Fabra, 2018; Salega y Fabra, 2013) han sugerido que estos cambios climáticos pudieron haber afectado la disponibilidad de recursos y, de esa forma, pudieron haber incidido en las condiciones generales de salud, sometiendo a las poblaciones a niveles de estrés y a un posible déficit nutricional desde edades tempranas. Particularmente, la presencia de hipoplasias del esmalte dental sugiere que las poblaciones asentadas en el centro de Argentina sufrieron una disminución en los niveles de salud en momentos finales del Holoceno tardío (González, 2016): las elevadas prevalencias de esta patología de origen metabólico sugieren que los más afectados por episodios de estrés fueron individuos adultos jóvenes y medios en toda el área estudiada, siendo las mujeres las que presentan mayores prevalencias, particularmente en llanuras. Las causas que permiten explicar estos eventos de estrés en la población, afectando en mayor medida a individuos femeninos, pueden ser numerosas, entre ellas, causas culturales o sociales que incidieron en las condiciones de salud femenina durante la niñez. Sin embargo, consideramos posible que se haya producido una reducción en la disponibilidad de alimentos hacia fines del Holoceno tardío, y quizás una mayor competencia por los recursos, lo cual habría limitado la oferta alimenticia, o incluso habría aumentado el consumo de ciertos alimentos, derivando posiblemente en un estrés nutricional que afectó mayormente a los individuos femeninos.
Es sugerente pensar que las diferencias en la estatura entre individuos de ambos sexos, y dentro de los mismos sexos, en dos momentos temporales que coinciden con cambios climáticos marcados, pueden estar relacionados en algún punto, por ejemplo, con la relación que existe entre las condiciones climáticas y la distribución de recursos potencialmente consumibles. Este tipo de procesos podrían explicar en parte los cambios ocurridos en la talla, pero aun así habría que evaluar otros factores, como son los procesos socioeconómicos y los cambios en el acervo genético de las poblaciones.
Numerosos son los trabajos que abordan la relación entre los cambios en las condiciones socioeconómicas con respecto a la variación de la tendencia secular de la talla (Cardoso y Gomes, 2008; Cardoso, 2008; Gustafsson et al., 2007; Shin et al., 2012). En el caso de Sudamérica, Abarca Labra (2011) relaciona el incremento de la talla con la implementación de políticas sociales y económicas que favorecieron a sectores de bajos recursos.
Los trabajos bioarqueológicos sobre la dieta y la salud llevadas adelante por González (2016) y Fabra y González (2015) desde la antropología dental, han evidenciado que la moderada frecuencia de patologías de origen infeccioso y metabólico concuerdan con los valores esperados para poblaciones con economía cazadora-recolectora y de carácter mixto. El análisis de isotopos estables complementa la información brindada por los indicadores dentales: los valores más positivos de 13Ccol en el período posterior a 1200 años AP sugieren que los individuos femeninos consumían alimentos producidos, en mayor cantidad que sus pares masculinos, quienes, por otro lado, según la información isotópica brindada por el nitrógeno 15 (15N), habrían incorporado a su dieta mayor cantidad de proteínas animales, sobre todo en llanuras.
La producción de alimentos implicó un cambio en las actividades cotidianas realizadas por las personas y provoco modificaciones en el modo de vida de los individuos, observándose un leve deterioro en la salud para momentos tardíos del Holoceno (González, 2016). Estas modificaciones en los modos de vida en los siglos previos a la conquista española pueden estar relacionadas no solo con los cambios ya mencionados, sino también con las mayores exigencias que implicaron la obtención de alimentos en un contexto de reducción de la oferta ambiental. Esta reducción en la cantidad y/o variedad de recursos sería producto de la intensificación en la ocupación de espacios por el crecimiento poblacional y de la búsqueda de nuevos territorios, así como por cambios ambientales (Laguens y Bonnin, 2009).
Por otro lado, la presencia de hipoplasias en el esmalte dental sugiere que las poblaciones sufrieron una disminución en los niveles de salud en momentos finales del Holoceno tardío. Las causas que permiten explicar estos eventos de estrés, que afectaron en mayor medida a individuos femeninos, pueden ser numerosas, entre ellas, causas culturales o sociales. Pero también se considera posible que se haya producido una reducción en la disponibilidad de alimentos hacia fines del Holoceno tardío como se planteó al tratar la influencia de factores climáticos, y quizás una mayor competencia por los recursos, lo cual habría limitado la oferta alimenticia, o incluso habría aumentado el consumo de ciertos alimentos. Todo esto derivó en un estrés nutricional, que afectó mayormente a los individuos femeninos (González, 2016: 269, 270) y su talla final.
En esta región se han llevado a cabo estudios sobre cambios degenerativos y entesiales en poblaciones de las sierras y las llanuras de Córdoba, a lo largo del Holoceno tardío. Salega (2016) y Salega y Fabra (2013) sugieren que, para momentos del Holoceno tardío final, disminuyen las frecuencias de modificaciones óseas en miembros inferiores y columna, a la vez que la prevalencia de cambios entesiales es mayor en miembros superiores. Además, observan una disminución en la frecuencia de facetas extra y extensiones articulares vinculadas a la locomoción y las posturas arrodilladas. Las autoras relacionan estos cambios con una reducción de la movilidad residencial y logística, como consecuencia de un aumento de la sedentarización hacia finales del Holoceno tardío.
Con respecto a la procedencia geográfica se observa una mayor frecuencia de cambios degenerativos en los individuos de llanuras, con mayor cantidad de porciones anatómicas comprometidas (tanto en columna vertebral como en esqueleto apendicular), y mayor grado de desarrollo y extensión, lo cual podría sugerir un mayor desgaste de las articulaciones por actividades físicas más demandantes con respecto a los individuos que habitaban en la zona de sierras.
Allen (1984) propone que la intensidad y la exigencia de las actividades físicas cotidianas, en especial aquellas que son iniciadas en la niñez y poseen continuidad durante la fase de crecimiento, pueden repercutir en la talla final de los individuos, presentándose disminuciones en la estatura (Allen, 1984). En la región estudiada, la exigencia y la intensidad de las tareas diarias no habría repercutido directamente sobre la estatura de los individuos masculinos, pues se observa mantenimiento de la talla a lo largo del Holoceno tardío. Para el caso de los individuos femeninos, tampoco se daría una correlación entre exigencia física cotidiana y talla, ya que en momentos iniciales del Holoceno tardío se ven tallas elevadas y frecuencias más altas para cambios degenerativos y entesiales (Salega, 2016). Para el caso de la procedencia geográfica, los individuos masculinos poseen mayor estatura en sierras, posiblemente favorecidos por los microambientes, propios de los valles serranos, y por las menores distancias recorridas para el abastecimiento de recursos, lo que implica menores índices de actividad física y bajas posibilidades de que afecte la estatura final de los individuos. En el caso de las llanuras, los individuos femeninos poseen mayor talla y es en esta región en donde se revelaron las mayores prevalencias de cambios degenerativos y entesiales (Salega, 2016; Salega y Fabra, 2013).
Los resultados observados en nuestro trabajo coinciden parcialmente con los obtenidos por Millán y colaboradores (2013), quienes observaron un mantenimiento a lo largo del período estudiado en la talla tanto en individuos femeninos como masculinos, con un leve incremento en la estatura de los segundos, y un incremento en el dimorfismo sexual. Las autoras sugieren que el incremento en el dimorfismo sexual estaría relacionado con una mejora en las condiciones generales de vida en momentos tardíos. A diferencia de lo ocurrido en las poblaciones patagónicas, en la región estudiada se observó un decrecimiento para las mujeres. En general, los individuos masculinos son los más vulnerables a posibles disrupciones nutricionales y/o ambientales, ya que los individuos femeninos se encuentran rigurosamente encaminados en su canal de crecimiento, característica que Waddington (1970) ha llamado “la mejor canalización femenina”, la cual confiere mayor ecoresistencia o vulnerabilidad ambiental acotada. Esta mejor canalización femenina significa que, en situaciones en que las poblaciones se ven afectadas por condiciones desfavorables, el crecimiento femenino se encuentra rigurosamente encaminado y tiene mayores posibilidades de expresar su talla total (Lieberman, 1982). ¿Cómo podemos explicar entonces la disminución en la talla en individuos femeninos? ¿Qué otros procesos o factores pueden ser los causantes?
El último factor que considerar al momento de explicar variaciones en la talla en las sociedades que habitaron las Sierras Pampeanas es el impacto que podrían haber tenido los cambios en el acervo genético de una población debido a distintos factores microevolutivos, por ejemplo, migraciones o flujo génico. En esta región es interesante evaluar si los cambios detectados en la composición genética de la población que habitó tanto las sierras como las llanuras de la provincia (Nores y Demarchi, 2011; Nores et al., 2011), pueden estar relacionados con las variaciones en la talla, a lo largo del tiempo y entre sexos.
Las investigaciones realizadas sobre ADN mitocondrial que partieron de un conjunto de muestras arqueológicas de sitios de la actual provincia de Córdoba sugieren que hacia 1200 años AP habría ocurrido una diferenciación genética entre poblaciones que habitaron distintos espacios geográficos (Nores y Demarchi, 2011; Nores et al., 2011; Fabra et al., 2014): en el Holoceno tardío inicial los haplogrupos con mayor prevalencia son el C en la región serrana, y el C y el D en la región de llanuras. Luego, se observa un cambio en la frecuencia de haplogrupos en cada región, siendo los haplogrupos B y C los más frecuentes en sierras, y A, C y D en llanuras. Existe una alta frecuencia del haplogrupo C en toda la muestra de Sierras Centrales.
Estos resultados sugieren un aporte inmigratorio diferente hacia las dos subregiones a partir de 1200 años AP, representado por presencia del haplogrupo B para sierras y del A para llanuras. La presencia del haplogrupo A sugiere flujo génico desde el este y el noreste, de donde provienen grupos guaraníes. Para el caso del B, aporte inmigratorio de regiones andinas y/o chaqueñas. La presencia de haplogrupos C y D hace suponer vínculos con poblaciones patagónicas (Nores y Demarchi, 2011; Nores et al., 2011). Los estudios de ADN mitocondrial también detectaron diferencias significativas entre las muestras arqueológicas y las de poblaciones contemporáneas (Nores y Demarchi, 2011). No hay que dejar de lado que se dieron determinadas continuidades de tipo genético a lo largo del tiempo, como en el haplogrupo D1j, caracterizado por su alta frecuencia y posible origen en la región (García et al., 2012).
En un estudio llevado adelante por Grasgruber y colaboradores (2014, 2017) para poblaciones del este europeo (Bosnia-Herzegovina), se asocian las altas tallas con la distribución de un linaje genético masculino: el haplogrupo Y I-170. Según estos autores, las condiciones socioeconómicas y nutricionales no son consideradas óptimas para la región, en comparación con países industrializados del oeste los cuales también poseen tallas promedio altas, producto de los beneficios positivos del modelo industrial, donde se da lugar a la explicación del potencial genético para esta región bajo estudio.
Para el caso de la Argentina hay pocos antecedentes de investigaciones que vinculan la tendencia secular de la talla con el aporte que puede realizar la genética, como factor explicativo. Berón (2007) menciona cómo la llegada de poblaciones foráneas a la región del sur de La Pampa para periodos tardíos, proceso conocido como araucanización, generó un cambio en la talla, siendo la población migrante de menor estatura que las poblaciones Tehuelches que habitaban la región. Se puede pensar que la talla de los individuos de Sierras Centrales pudo verse influenciada por el ingreso de poblaciones alrededor de 1200 años AP, que se vería reflejado en los cambios de estaturas para este periodo, junto a la variación secular negativa detectada en individuos femeninos y una amplia representación de tallas en la muestra estudiada.
A modo de conclusión, los estudios arqueológicos y bioantropológicos sobre poblaciones que habitaron la región central de Argentina dan cuenta de una historia biológica compleja, cuya variabilidad fue moldeada fundamentalmente por el flujo génico como fuerza evolutiva preponderante (Fabra, 2009, 2009b, 2014; Fabra y Demarchi, 2009, 2011, 2012a, 2012b; Fabra et al., 2012, 2014; Laguens et al., 2007). ¿Cómo vincular los cambios temporales y espaciales en la talla, observados en el presente estudio, con la variabilidad biológica, y los cambios y continuidades en los modos de vida de las poblaciones? La amplia distribución de estaturas a lo largo del Holoceno tardío sugiere que no podemos explicar las variaciones observadas en distintas regiones y por sexo, atendiendo a un único factor explicativo. Más bien, resulta interesante pensar en causas múltiples que están interviniendo en los procesos que modelan la talla de los individuos a lo largo del tiempo.
Los procesos de cambio social y económico iniciados entre los 2000 y 1500 años AP habrían llevado a un desmejoramiento de la salud general de las poblaciones de Sierras Centrales; sin embargo, no habrían impactado de forma homogénea en toda la población, ya que los individuos masculinos mantuvieron y hasta incrementaron levemente su talla. Consideramos posible que se haya producido una reducción en la disponibilidad de alimentos hacia fines del Holoceno tardío, y quizás una mayor competencia por los recursos, lo cual habría limitado la oferta alimenticia o incluso habría aumentado el consumo de ciertos alimentos, derivando en un estrés nutricional, que afectó mayormente a los individuos femeninos e impactó en su talla. En el caso puntual de los individuos femeninos, si bien la presencia de mayor frecuencia de indicadores de estrés metabólico sugiere dichos procesos (González, 2016; Fabra y González, 2015), estos no habrían sido ni tan marcados ni tan extendidos en la población como para explicar una disminución en la talla. No podemos dejar de pensar que el cambio en la composición genética evidenciado alrededor de 1200 años AP puede haber generado, en parte, el patrón observado de variación en la talla. Ahondar en los estudios bioarqueológicos y arqueogenéticos de las poblaciones de ambas regiones puede otorgar nuevos datos para evaluar qué variables están interviniendo en los cambios de la talla durante el Holoceno.
Agradecimientos
Los autores desean agradecer al Museo de Antropología (FFyH, UNC), al Museo Histórico Municipal de La Para (La Para, Córdoba), al Museo de la región de Ansenuza “Aníbal Montes” (Miramar, Córdoba), al Museo Arqueológico Provincial “Aníbal Montes” (Río Segundo, Córdoba), al Museo Comechingón (Mina Clavero, Córdoba) y al Museo Capitán Juan de Zevallos (Valle Hermoso, Córdoba), por el acceso brindado para el estudio de las colecciones bioarqueológicas. Muy especialmente agradecen al Dr. Dario Demarchi por su asesoramiento para la realización de los análisis estadísticos. A los evaluadores/as anónimos/as por sus valiosos comentarios. Este trabajo se desarrolló en el marco de los proyectos PICT 2013-2028, 2015-3155 y SECyT-UNC (2014-2015).
Referencias bibliográficas
Abarca Labra, V. (2011). Efectos de la nutrición sobre el Dimorfismo Sexual expresado en la Estatura (SSD) de una muestra de población Chilena Subactual. Memoria para optar por el Título Profesional de Antropología Física. Universidad de Chile, Facultad de Ciencias Sociales, Departamento de Antropología.
Allen, L.H. (1984). Functional indicators of nutritional status of the whole individual or the community. Clinical Nutrition 3 , 169 - 75.
Beguelin, M. (2009). Variación Geográfica en la Morfología del Esqueleto Postcraneal de las Poblaciones Humanas de Pampa y Patagonia durante el Holoceno Tardo: Una Aproximación Morfométrica. PhD dissertation. Universidad Nacional de La Plata, Argentina.
_____. (2010). Tamaño corporal y temperatura ambiental en poblaciones cazadoras recolectoras del Holoceno tardío de Pampa y Patagonia. Revista Argentina de Antropología Biológica, 12 (1), 27 - 36.
_____. (2011). Stature estimation in a Central Patagonian prehispanic population: development of new models considering specific body proportions. International Journal of Osteoarchaeology, DOI: 10.1002/oa.1117
Beguelin, M., y Barrientos, G. (2005). Variación morfométrica postcraneal en muestras tardías de restos humanos de Patagonia: una aproximación biogeográfica. Intersecciones en Antropología, 7, 49 - 62.
Beguelin, M., y Gonzalez, P. (2008). Estimación del sexo en poblaciones del Sur de Sudamérica mediante funciones discriminantes para el fémur. Revista Argentina de Antropología Biológica, 10 (2), 55 - 70.
Bernal, V., Beguelin, M., Gordon, F., Cobos, V., González, P.N., & Lotto, F. (2013). Craniofacial variation, body size and ecological factors in aboriginal populations from central Patagonia (2000-200 years BP). Homo– J.Comp.Hum.Biol . 65 (2).
Berón, M.A. (2007). Integración de evidencias para evaluar la dinámica y circulación de poblaciones en las fronteras del río Colorado. En F. Morello, M. Martinic, A. Prieto y G. Bahamondes (eds.), Arqueología de Fuego Patagonia. Levantando piedras, desenterrando huesos y develando arcanos: 173 - 188. Punta Arenas, Ediciones CEQUA.
Bogin, B. (1999). Patterns of human growth. Second edition. Cambridge, Cambridge University.
Bonifligio, M. (2009). Arqueología de la zona lacustre de Córdoba (N.E.). Un contexto para armar. Actas Séptimas Jornadas de Arqueología y Etnohistoria del Centro Oeste Argentino [pp. 83 - 101]. Universidad Nacional de Río Cuarto.
Buikstra, J., & Ubelaker D. (Eds.). (1994). Standards for Data Collection from Human Skeletal Remains. Arkansas Archaeological Survey Research Series (44).
Cardoso, H.F.V. (2008). Secular changes in body height and weight of portuguese boys over one century. American Journal of Human Biology 20, 270 – 277.
Cardoso, H.F.V., & Gomes J.E.A. (2009). Trends in adult stature of peoples who inhabited the modern Portuguese territory from the Mesolithic to the late 20th century. International Journal of Osteoarchaeology, 19(6), 711– 725.
Carignano , C. (1999). Late Pleistocene to recent climate change ln Córdoba Province, Argentina: Geomorphological evidence. Quaternary International, 57/58, 117-134.
Cohen, M.N., & G.J, Armelagos. (1984). Paleopathology at the Origins of Agriculture. Editors Orlando, FL: Academic Press.
Coltrain, J., & Leavitt, S. (2002). Climate and diet in Fremont prehistory: Economic variability and abandonment of maize agriculture in the Great Salt Lake basin. American Antiquity 67, 453 - 485.
Fabra, M. (2008). Patrones geográficos de diferenciación craneofacial entre poblaciones de la región central del país y otras de Argentina: aportes desde la genética del paisaje. Revista del Museo de Antropología, 1(1): 13-24
_____ (2009). Historia de las poblaciones prehispánicas del sector austral de las Sierras Pampeanas: variabilidad morfológica y modelos arqueológicos. Tesis Doctoral. Facultad de Filosofía y Humanidades. Universidad Nacional de Córdoba. Ms.
_____. (2013). Variación epigenética craneofacial y dinámica evolutiva de poblaciones humanas del norte de la región pampeana durante el holoceno tardío. Revista Cuadernos del Instituto Nacional de Pensamiento Latinoamericano edición especial I Congreso Internacional de Arqueología de la Cuenca del Plata, 1 (1), 74 - 86.
_____. (2014). Historia de las poblaciones prehispánicas del sector austral de las Sierras Pampeanas: variabilidad morfológica y modelos arqueológicos. E-Book. Publicaciones de la Secretaria de Ciencia y Tecnología, FFyH, UNC. ISBN 978-950-33-1098-4
Fabra, M., Laguens, A., y Demarchi, D. (2005). Análisis intra e inter poblacional de rasgos craneanos no métricos en aborígenes prehispánicos del actual territorio de Córdoba. Revista Argentina de Antropología Biológica, 7(2), 47 - 65.
Fabra, M., y Demarchi, D. (2009). Variabilidad craneofacial en poblaciones del sector austral de las Sierras Pampeanas: aportes desde la morfometría geométrica. Revista Relaciones de la Sociedad Argentina de Antropología, 34, 1 - 24. ISSN 0325-2221.
_____. (2012). Variación Morfológica craneofacial y estructura genética de poblaciones del centro de Argentina. Revista Argentina de Antropología Biológica, 14(1), 45 - 56.
_____. (2013). Análisis morfogeométrico aplicado al estudio de los patrones espaciales y temporales de variación morfológica craneofacial en poblaciones del centro de Argentina. Revista Cuadernos del INAPL, edición especial I Congreso Internacional de Arqueología de la Cuenca del Plata 1(1), 87 - 101.
Fabra,M. Salega, M.S., y Gonzalez, C.V. (2009). Comportamiento mortuorio en poblaciones prehispánicas de la región austral de las Sierras Pampeanas durante el Holoceno. Revista Arqueología 15, 165 – 186.
Fabra M., González C.V., y Salega, M.S. (2012). Modos de vida e historia biológica de poblaciones de las Sierras y las Llanuras de Córdoba (Argentina): Aproximaciones desde el Registro Biorqueológico. Revista Argentina de Antropología Biológica. 14(Número Especial), 87 - 104.
Fabra, M., Nores, R., Salega, S., y González, CV. (2014). Entre las sierras y el Mar: investigaciones bioarqueológicas en el noroeste de la región pampeana (costa sur Laguna Mar Chiquita, Córdoba, Argentina). En Luna, L. Aranda, C. y J. Suby (eds.). Avances recientes en la Bioarqueología Sudamericana, [pp. 205-230], Grupo de Investigación en Bioarqueología, ISBN 978-987-27997-1-7.
Fabra, M., y González, CV. (2008). Análisis de bioindicadores dietarios en poblaciones prehispánicas del Centro de Argentina en el Holoceno Tardío. ArqueoWeb, Revista sobre Arqueología en Internet , 10 (1): 1-12.
_____. (2015). Diet and oral health of population that inhabited central Argentina (Córdoba province) during late Holocene. International Journal of Osteoarchaeology. DOI: 10.1002/oa.2272, 25: 16- 175.
Formicola, V., & Giannecchini, M. (1999). Evolutionary trends of stature in Upper Paleolithic and Mesolithic Europe. Journal of Human Evolution, 36(3), 339 - 33.
García, A., y Demarchi, D. (2006). Linajes parentales amerindios en poblaciones del Norte de Córdoba. Revista Argentina de Antropología Biológica 8(1), 57 - 71.
García, A., Pauro, M., Nores, R., Bravi, C.M., & Demarchi, D. (2012). Phylogeography of mitochondrial haplogroup D1: An early spread of subhaplogroup D1j from Central Argentina. American Journal of Physical Anthropology 149, 583 - 590.
González, C.V. (2016). Dieta y salud oral en poblaciones del centro de la Argentina durante el Holoceno Tardío. Tesis doctoral Inédita, FFyH, UNC.
González, C.V., y Fabra, M. (2018). Desgaste dental y hábitos dietarios en poblaciones arqueológicas del centro de Argentina. Arqueología 24(2), 133 - 159.
Grasgruber, P., Cacek, J., Kalina, T., & Sebera, M. (2014). The role of nutrition and genetics as key determinants of the positive height trend. Economics and Human Biology 15, 81 - 100.
Grasgruber, P., Popović, S., Bokuvka, D., Davidović, I., Hřebíčková, S., Ingrová, P., Potpara, P., Prce, S., & Stračárová, N. (2017). The mountains of giants: an anthropometric survey of male youths in Bosnia and Herzegovina. R. Soc. Open Sci. 4: 161054. http://dx.doi.org/10.1098/rsos.161054
Gustafsson, A., Wenderlin, L., Tullberg, B., & Linderfors, P. (2007). Stature and Sexual Stature Dimorphism in Sweden from the 10th to the end of the 20th Century. American Journal of Human Biology 19, 861 - 870.
Kemkes-Grottenthaler, A. (2005). The short die young: the interrelationship between stature and longevity-evidence from skeletal remains. American Journal of Physical Anthropology 128(2), 340 - 347.
Hernández, M., García-Moro, C., & Lalueza-Fox, C. (1998). Stature Estimation in Extinct Aónikenk and the Myth of Patagonian Gigantism. American Journal of Physical Anthropology 105, 545 - 551
Koepke, N. (2016). The biological standard of living in Europe from the late iron age to the little ice age. En: Komlos, J. y Kelly, I. The Oxford Handbook of Economics and Human Biology [pp. 70 - 109]. Oxford: Oxford University Press.
Komlos, J. (1998). Shrinking in a Growing Economy. The Mystery of Physical Stature during the Industrial Revolution. Journal of Economic History 58, 779 - 802.
Krenzer, U. (2006). Compendio de métodos antropológico-forenses para la reconstrucción del perfil osteo-biológico. Serie de Antropología Forense, Centro de análisis forense y ciencias aplicadas, Guatemala, ISBN 99922-859-2-3.
Laguens, A. (1999). Arqueología del contacto hispano-indígena. Un estudio de cambios y continuidades en las Sierras Centrales de Argentina. BAR International Series 801. Oxford.
Laguens, A., Fabra, M., Dos Santos, G.M., & Demarchi, D. (2009). Paleodietary inferences based on isotopic evidences for populations of the Central Mountains of Argentina during the Holocene. International Journal of Osteoarchaeology 19, 237 - 249.
Laguens , A., y Bonnin, M. (2009). Sociedades indígenas de las Sierras Centrales. Arqueología de Córdoba y San Luis. Editorial Universidad Nacional de Córdoba, Córdoba.
Lieberman, L. (1982). Normal and abnormal sexual dimorphic pattern of growth and development. En Hall R (Ed). Sexual Dimorphism in Homo sapiens [pp. 253-316] New York: Praeger.
Martin, R., & Saller, K. (1957). Lehrbuch der Anthropologie. En Systematischer Darstellumg mit besonjerer Beruckichtigung der Anthropologischen methodes. Band I. Stuttgart, Gustav Fischer Verlag.
Martínez Sarasola, C. (1992). Nuestros paisanos los indios. “Vida, historia y destino de las comunidades indígenas en la Argentina”. Buenos Aires: Emecé Editores.
Medina M., Pastor S., & Recalde A. (2016). The archaeological landscape of Late Prehispanic mixed foraging and cultivation economy (Sierras of Cordoba, Argentina). Journal of Anthropological Archaeology 42, 88 - 104.
Millán, A.G., Gomez Otero, J., y Dahinten, S. (2013). Tendencia secular de la estatura en poblaciones humanas del Valle inferior del Rio Chubut y de la costa centro-septentrional (Patagonia) durante el Holoceno tardío. Relaciones de la Sociedad Argentina de Antropología 38(2), 421 - 440.
Neves, W. A., & Costa, M.A. (1998). Adult stature and standard of living in the prehistoric Atacama desert. Current Anthropology 39 (2), 278 - 281.
Nores, R., y Demarchi, D. (2011). Análisis de haplogrupos mitocondriales en restos humanos de sitios arqueológicos de la provincia de Córdoba. Revista Argentina de Antropología Biológica, 13 (1), 43 - 54.
Nores, R., Fabra, M., y Demarchi, D. (2011). Variación temporal y espacial en poblaciones prehispánicas de Córdoba. Análisis de ADN antiguo. Revista del Museo de Antropología 4, 187 - 194.
Nores, R., Fabra, M., García, G., y Demarchi, D. (2017). Diversidad genética en restos óseos humanos arqueológicos del sitio El Diquecito (costa sur, laguna Mar Chiquita). Revista Argentina de Antropología Biológica. 19 (1), 1 – 12. DOI 10.17139
Pauro , M., García, A., Bravi, C., y Demarchi, D. (2010). Distribución de haplogrupos mitocondriales alóctonos en poblaciones rurales de Córdoba y San Luis. Revista Argentina de Antropología Biológica 12(1), 47 - 55.
Piovano, E.L., Ariztegui, D. Córdoba, F., Cioccale, M., & Sylvestre, F. (2009). Hydrological Variability in South America Below the Tropic of Capricorn (Pampas and Patagonia, Argentina) During the Last 13.0 Ka. En Vimeux F., Sylvestre, F. y Khodri, M. (Eds.). Past Climate Variability in South America and Surrounding Regions [pp. 323- 351]. France: Springer.
Pomeroy, E., & Stock, J.T. (2012). Estimation of stature and body mass from the skeleton among coastal and mid altitude Andean populations. American Journal of Physical Anthropology, 147, 264 - 279.
Roosevelt, A.C. (1984). Population, health and the evolution of subsistence: conclusions from the conference. En Cohern M.N. y Armelagos G.J. (Eds). Paleothology at the origins of agriculture [pp. 271 – 305]. Orlando: Academic Press.
Ruff C.B., Scott W., & Liu, A. (1991). Articular and diaphyseal remodeling of the proximal femur with changes in body mass in adults. American Journal of Physical Anthropology, 86, 397 - 413.
Salega, MS., y Fabra, M. (2013). Niveles de actividad física en poblaciones de las sierras y las llanuras de la provincia de Córdoba (Argentina) durante el Holoceno tardío. Relaciones de la Sociedad Argentina de Antropología, 38(2), 401 - 420.
Salega, M.S. (2016). Prácticas cotidianas, niveles de actividad física y modos de vida en poblaciones del sector austral de las Sierras Pampeanas durante el Holoceno tardío, tesis doctoral Inédita, Facultad de Filosofía y Humanidades, Universidad Nacional de Córdoba
Scheuer L., & Black, S. (2000). Developmental juvenile osteology. Academic Press, San Diego, New York, Tokyo.
Shin, D.H., Oh, C.S., Kim, Y.S., & Hwang, Y.I. (2012). Ancient-to-modern secular changes in Korean stature. American Journal of Physical Anthropology, 147, 433– 442.
Steele, D.G., & Mckern, T.W. (1969). A method for assessment of Maximum Long Bone Length and Living Stature from fragmentary long bones. American Journal of Physical Anthropology, 31, 215 - 227.
Tanner, J.M. (1962). Growth at adolescence (2da Ed). Oxford, Blackwell Scientific publications, and Springfield Thomas.
Trotter, M., & Gleser, G.C. (1958). A re- evaluation of estimation of stature base on measurements of stature taken during life and long bones after death. American Journal of Physical Anthropology 16, 79 - 123.
Van Wieringen, J. (1986). Secular growth changes. En Flakner, F. y Tanner, J.M. (Eds). Human growth- A comprehensive Treatuse 3. New York: Plenum Press.
Waddington, C.H (Ed). (1970). Towards to Theorical Biology: three drafts. Edinburgh: Edinburgh University Press.
Notas